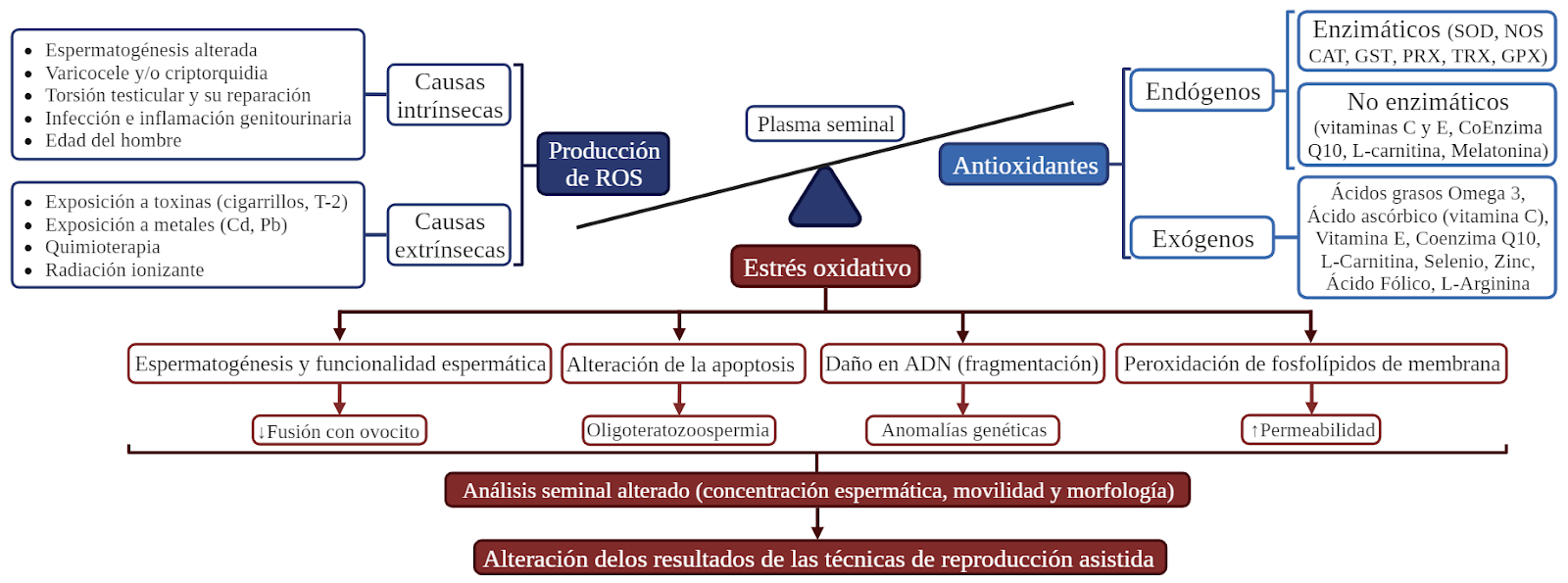
Graphical abstract. Causas del estrés oxidativo en el semen y consecuencias sobre los espermatozoides humanos y la fertilidad masculina.
1. ROS Y ESTRÉS OXIDATIVO
La infertilidad afecta al 17.5% de las parejas en edad reproductiva (OMS, 2023) y el factor masculino es la única causa en el 25% de los casos por una disminución de la calidad seminal de origen multifactorial (ambiental, genética, fisiológica, nutricional, etc.) (Auger et al., 2001), principalmente por el estrés oxidativo (30-80% de hombres infértiles) originado por elevadas concentraciones de especies reactivas de oxígeno (ROS) (Ko et al., 2014; Makker et al., 2009).
El oxígeno es un elemento necesario para la supervivencia celular, pero como resultado de los procesos fisiológicos y del metabolismo celular puede dar lugar a la formación de radicales libres de oxígeno (ROS). Su forma primaria es el anión superóxido (O2-), que surge de la adición de un electrón al O2 y puede convertirse en otras ROS como el peróxido de hidrógeno (H2O2), el radical hidroxilo (OH-), y el peroxilo (HO2). La presencia de electrones desapareados permite su interacción con aminoácidos, lípidos y ácidos nucleicos, provocando reacciones en cadena comprometen la vitalidad de la célula (Agarwal et al., 2014).
Se requieren niveles fisiológicos de ROS para el mantenimiento celular debido a su implicación en diversos procesos fisiológicos como son la maduración, hiperactivación y capacitación espermática, así como para la quimiotaxis, la unión a la zona pelúcida, la reacción acrosómica y la fusión entre ovocito y espermatozoide. A nivel testicular, las ROS son intermediarias implicadas en el tono vascular y la regulación génica (De Lamirande et al., 2009; Agarwal et al., 2014). En el semen, los principales productores de ROS son los espermatozoides inmaduros y los leucocitos, especialmente los neutrófilos. Ambas células aumentan su actividad mitocondrial y, adicionalmente, la vía de las NADHP, resultando en la producción de ROS (Nicolich et al., 2013).
En condiciones normales, existe un equilibrio entre la producción de ROS y la maquinaria antioxidante seminal y espermática (Ford et al., 2004). Sin embargo, este sistema puede desequilibrarse a favor de las ROS, ya sea por un aumento en su producción, por una deficiencia en la capacidad antioxidante o por ambas causas, generando un estado de estrés oxidativo presente en más del 80% de los hombres infértiles (Agarwal et al., 2011). Este estrés oxidativo puede generar daño espermático (Sikka et al., 2004) por inducción de peroxidación lipídica, incremento en la fragmentación de ADN (Moustafa et al., 2004), disminución en la movilidad y morfología espermática, y aumento de los fenómenos de apoptosis.
Actualmente, existen métodos comerciales capaces de medir el estrés oxidativo como el Kit Oxisperm, el cual se basa en el uso de un gel que reacciona con el anión superóxido (principal ROS responsable del estrés oxidativo en el semen humano), dando lugar a un precipitado que vira de color rosa a negro a concentraciones crecientes del anión superóxido. Por otra parte, este test sólo debe realizarse en muestras frescas ya que el proceso de criopreservación espermática puede inducir un incremento en los niveles de ROS y modificar el resultado del análisis. (Nicolich et al., 2013).
El aumento de estrés oxidativo espermático puede deberse a causas intrínsecas o extrínsecas (Cocuzza et al., 2007). Entre las causas intrínsecas se encuentran fallos en la espermatogénesis como la remodelación de la membrana del espermatozoide, las anomalías en la unión cabeza-cola, el desarrollo incompleto del acrosoma o las alteraciones en el citoesqueleto del espermatozoide (Sharma et al., 1996; Kothari et al., 2010). A nivel testicular existen diversas etiologías que aumentan la concentración de ROS por influjo de leucocitos como infecciones e inflamaciones genitourinarias o por aumento de la temperatura testicular y escrotal como en el varicocele (presente en el 30% de los hombres infértiles), la criptorquidia, la torsión testicular y su reparación. Por otra parte, la edad del hombre es un factor importante a tener en cuenta por su implicación en la producción y acumulación de ROS y por la asociada disminución de la esteroidogénesis y de la capacidad tamponadora de las células de Leydig (Cao et al., 2004).
De forma extrínseca al organismo, existen factores medioambientales que pueden provocar un aumento en los niveles de ROS y estrés oxidativo y alterar la espermatogénesis (Agarwal et al., 2003). Entre estos factores se encuentran la quimioterapia, la radiación ionizante y la exposición a toxinas y contaminantes como el metoxietanol presente en pinturas, líquidos de freno y otros químicos industriales (Syed et al., 1998), el tolueno presente en disolventes (Homma-Takeda et al., 2002), metales pesados como el cadmio (Koizumi et al., 1992) o el plomo (Hsu et al., 1997), las toxinas generadas al fumar cigarrillos (Mattison et al., 1982). Asimismo, estudios recientes (He et al., 2023) han demostrado que la toxina T-2 o micotoxina, producida por hongos y omnipresente en los cultivos agrícolas, alteraba significativamente la morfología de los testículos y de los espermatozoides en ratón y disminuía el nivel de testosterona y la concentración espermática. La ferroptosis inducida por la toxina T-2 podría ser mitigada eficazmente mediante la administración del inhibidor de la ferroptosis liproxstatina-1 o el antioxidante resveratrol, que proporciona un posible efecto terapéutico para el deterioro testicular.
Como se dijo anteriormente, el estrés oxidativo afecta a la espermatogénesis y a la funcionalidad espermática. En consecuencia, se alteran todos los parámetros del análisis seminal, incluyendo la concentración espermática (Agarwal et al., 2014), la movilidad y morfología (Aziz et al., 2004), la apoptosis celular (Agarwal et al., 2005) y la fragmentación del ADN espermático, lo que resulta en una disminución de la capacidad fecundante y peor desarrollo embrionario, con mayores tasas de aborto espontáneo y de recién nacidos enfermos por anomalías genéticas. Este efecto negativo se debe en gran parte a la alta susceptibilidad de las membranas espermáticas a la peroxidación por ROS por su elevada concentración de lípidos poliinsaturados El aumento de la peroxidación de fosfolípidos de membrana (Griveau et al. 1995) se traduce en la pérdida de integridad y fluidez de la misma, aumentando su permeabilidad y afectando a la movilidad espermática. Por otra parte, el estrés oxidativo aumenta la apoptosis en espermatozoides maduros, resultando en oligozoospermia (Gandini et al., 2000).
Todos estos efectos negativos sobre el espermatozoide pueden afectar a los resultados de las técnicas de reproducción asistida (TRA) (Agarwal et al., 2012)
2. ANTIOXIDANTES
Un antioxidante es una molécula capaz de retardar o prevenir la oxidación de otras moléculas evitando la generación de ROS y el daño celular. En un eyaculado se encuentran antioxidantes en el plasma seminal y, en menor medida, en los espermatozoides debido a su escaso contenido citoplasmático. Los antioxidantes provienen de dos vías, fisiológica y exógena.
Los antioxidantes endógenos que se encuentran en el semen se clasifican en enzimáticos (catalasa, glutatión peroxidasa y superóxido dismutasa) y no enzimáticos (carnitina, carotenoides, glutatión, urato y vitaminas C y E).
Los antioxidantes enzimáticos actúan a nivel de metabolismo directo de ROS, de la síntesis y reducción del glutatión, ciclos redox de tiol y activación de la expresión de algunas de estas enzimas (Drummond et al., 2000; Yu et al., 2015). La mayoría están reguladas por el factor de transcripción nuclear NRF2 mediante el elemento de respuesta antioxidante (ARE) localizado en los promotores de los principales genes del sistema antioxidante en semen. En condiciones normales, NRF2 se une a la proteína KEAP1 identificativa para la degradación. El estrés oxidativo disocia esta unión, permitiendo la translocación de NRF2 al núcleo y su unión a los ARE, que activa la expresión de genes que codifican para enzimas antioxidantes. Entre las enzimas del sistema antioxidante endógeno del semen que están bajo regulación de NRF2 se encuentra el conjunto de formas de la superóxido dismutasa (SOD), que cataliza la dismutación del radical superóxido a peróxido de hidrógeno fundamentalmente en el epidídimo, protegiendo de daños provocados por un exceso de O2 y aumentando los niveles de H2O2 en la eyaculación que estimula la cascada de reacciones de hiperactivación y capacitación del espermatozoide (Lifeng et al., 2014). La catalasa (CAT) cataliza la descomposición del peróxido de hidrógeno en oxígeno y agua en la mayoría de tejidos del tracto genital masculino, resultando esencial ante concentraciones elevadas de H2O2. La óxido nítrico sintasa (NOS) transforma la L-arginina en óxido nítrico (NO), que está relacionado con la función eréctil, la movilidad espermática y la reacción acrosómica, y actúa como antioxidante al evitar la peroxidación de lípidos de membrana (Drummond et al., 2000). La glutatión S-transferasa (GST) inactiva los agentes quimioterapéuticos productores de ROS. Otras enzimas que neutralizan las ROS son las peroxiredoxinas (PRX) y las tiorredoxinas (TRX), moduladas por la glutatión peroxidasa (GPX). Entre el conjunto de moléculas antioxidantes no-enzimáticas del semen se encuentran la vitamina E, la vitamina C, la coenzima Q10, la L-carnitina y la melatonina.
Por otra parte, los antioxidantes exógenos se obtienen a través de la ingesta de alimentos (frutas, verduras, frutos secos) o de suplementos nutricionales (suplementos vitamínicos, minerales o plantas medicinales), pudiendo mejorar la calidad seminal (Bermejo et al., 2014).
Entre las principales moléculas estudiadas se encuentran los ácidos grasos Omega 3 como el ácido linolénico (AL), el ácido eicosapentanoico (EPA) y el ácido docosahexanoico (DHA), constituyentes de membranas celulares e indispensables en la capacidad fecundante de los espermatozoides. Safarinejad et al. (2011) evaluaron el efecto de la suplementación con ácidos grasos omega 3 (EPA+DHA) sobre la calidad seminal de pacientes oligoastenoteratozoospérmicos (OAT) y sobre la capacidad antioxidante del plasma seminal, mostrando una mejora significativa en la concentración, movilidad y morfología espermática, así como una mejora en la capacidad antioxidante evaluada mediante la cuantificación de niveles de SOD y CAT. Adicionalmente, Comhaire et al. (2000) evaluaron el efecto de la administración conjunta de antioxidantes (N-acetilcisteína o vitaminas A y E) con ácidos grasos esenciales (DHA + ácido gamma linolénico + ácido araquidónico) durante un periodo de 6 meses, que mejoraba la concentración espermática en pacientes oligozoospérmicos, pero no mejoraba su movilidad ni su morfología espermática.
Por otro lado, se estudia el efecto antioxidante de múltiples vitaminas. La Vitamina C o ácido ascórbico es el principal antioxidante presente en el plasma seminal con valores 10 veces superiores a los del plasma sanguíneo (Lewis et al., 1997). Dawson et al. (1997) administraron vitamina C a pacientes fumadores y observaron una mejora de su concentración, morfología y vitalidad espermáticas. Esta mejora de movilidad y morfología espermática coincide con la observada por Cyrus et al. (2015) al administrar Vitamina C durante tres meses a pacientes tras varicocelectomía. La vitamina E in vitro reduce la peroxidación lipídica, protege a los espermatozoides de una disminución en su movilidad (Askari et al.) y disminuye la producción de H2O2 y la fragmentación de ADN. Su administración prolongada mejoraba la movilidad espermática y disminuía los niveles de malonaldehído (marcador de peroxidación) en pacientes astenozoospérmicos (Suleiman et al., 1996). De acuerdo a Suleiman et al. (1996), la administración combinada de estas dos vitaminas durante dos meses en pacientes infértiles mejoraba la concentración espermática y disminuía la fragmentación del ADN. La administración de zinc, ya presente en el plasma seminal, mejora la concentración, la movilidad progresiva y la capacidad fecundante de los espermatozoides en pacientes astenozoospérmicos (Omu et al., 1998). Las vitaminas A, C y D potencian este efecto (Omu et al., 2008) y la acción antioxidante del selenio (Scott et al., 1998).
En la maduración espermática, un antioxidante implicado es la L-carnitina, cuya concentración se correlaciona con la concentración espermática, el porcentaje de espermatozoides con movilidad progresiva y el porcentaje de espermatozoides con morfología normal (Agarwall et al., 2004). Además, en combinación con L-arginina y ginseng durante tres meses, Morgante et al. (2010) observaron una mejora mayor de la movilidad espermática. Por otro lado, la coenzima Q10 posee propiedades antioxidantes por su implicación en la cadena transportadora de electrones, y su administración a diferentes concentraciones y durante diferentes periodos de administración parece incrementar la movilidad y concentración espermática (Balercia et al., 2009; Safarinejad et al., 2009). Finalmente, se han asociado niveles bajos de ácido fólico en el plasma seminal con mayores índices de fragmentación de ADN espermático por su implicación en la síntesis de ADN y ARN (Boxmeer et al., 2009). Su administración aislada no ha sido estudiada, pero en combinación con zinc durante 26 semanas aumentaba la concentración espermática (Ebisch et al., 2006), y en pacientes astenozoospérmicos tras una varicocelectomía mejoraba la calidad seminal (Azizollahi et al., 2013).
Además, se sigue analizando el efecto antioxidante de la administración de diferentes combinaciones de estas moléculas y su influencia sobre la calidad seminal. Algunos de estos ensayos quedan reflejados en la Tabla 1.
.")
Tabla 1. Resumen de resultados de distintos ensayos clínicos con antioxidantes (Martínez et al., 2017).
3. CONCLUSIÓN
En conclusión, teniendo en cuenta las consecuencias del estrés oxidativo en la función espermática y en las TRA, éste debería ser analizado de manera más rutinaria en las muestras seminales para clasificar correctamente la causa de infertilidad de los pacientes. De este modo, podría mejorarse la calidad espermática con suplementos nutricionales para, finalmente, aumentar las posibilidades de éxito de las TRA.
4. BIBLIOGRAFÍA
Agarwal A, Aponte-Mellado A, Premkumar BJ, Shaman A, Gupta S. The effects of oxidative stress on female reproduction: a review. Reprod Biol Endocrinol. 2012 Jun 29;10:49.
Agarwal A, Nallella KP, Allamaneni SS, Said TM. Role of antioxidants in treatment of male infertility: an overview of the literature. Reprod Biomed Online. 2004 Jun;8(6):616-27.
Agarwal A, Said TM. Oxidative stress, DNA damage and apoptosis in male infertility: a clinical approach. BJU Int. 2005 Mar;95(4):503-7.
Agarwal A, Saleh RA, Bedaiwy MA. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil Steril. 2003 Apr;79(4):829-43.
Agarwal A, Sekhon LH. Oxidative stress and antioxidants for idiopathic oligoasthenoteratospermia: Is it justified? Indian J Urol. 2011 Jan;27(1):74-85.
Agarwal A, Sharma RK, Sharma R, Assidi M, Abuzenadah AM, Alshahrani S, et. al. Characterizing semen parameters and their association with reactive oxygen species in infertile men. Reprod Biol Endocrinol. 2014 May 7;12:33.
Agarwal A, Virk G, Ong C, du Plessis SS. Effect of oxidative stress on male reproduction. World J Mens Health. 2014 Apr;32(1):1-17.
Agarwall A, Tamer MS. Carnitines and male infertility. Reproductive Biomedicine Online (2004), 8(4):376-384
Aitken RJ, Clarkson JS, Fishel S. Generation of reactive oxygen species, lipid peroxidation, and human sperm function. Biol Reprod. 1989 Jul;41(1):183-97.
Aitken RJ, Koppers AJ. Apoptosis and DNA damage in human spermatozoa. Asian J Androl. 2011 Jan;13(1):36-42.
Aksoy Y, Aksoy H, AltInkaynak K, AydIn H R I , Özkan A. Sperm fatty acid composition in subfertile men. Prostaglandins, Leukotrienes and Essential Fatty Acids 2006; 75, 75-79.
Arduini A. Carnitine and its acyl esters as secondary antioxidants. AM Heart J (1992) 123:1726-7
Askari HA, Check JH, Peymer N, Bollendorf F. Effect of natural antioxidants tocopherol and ascorbics acids in maintenance of sperm activity during freeze thaw process. Archives of Andrology 33:11-15
Auger J, Eustache F, Andersen AG, Irvine DS, Jørgensen N, Skakkebaek NE, et al. Sperm morphological defects related to environment, lifestyle and medical history of 1001 male partners of pregnant women from four European cities. Hum Reprod. 2001Dec;16(12):2710-7.
Aziz N, Saleh RA, Sharma RK, Lewis-Jones I, Esfandiari N, Thomas AJ Jr, et al. Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil Steril. 2004 Feb;81(2):349-54.
Azizollahi G, Azizollahi S, Babaei H, Kianinejad M, Baneshi MR, NematollahiMahani SN. Effects of supplement therapy on sperm parameters, protamine content and acrosomal integrity of varicocelectomized subjects. J Assist Reprod Genet (2013) 30(4): 593-599
Balercia G, Buldreghini E, Vignini A, Tiano L, Paggi F, Amoroso S et al. Coenzyme Q10 treatment in infertile men with idiopathic asthenozoospermia: a placebo-controlled, double-blind randomized trial. Fertility and Sterility (2009), May 91(5): 1785-1792
Bermejo P, Cuadrado I, Ortega JC, De las Heras B. Evaluación de la actividad antioxidante de suplementos dietéticos utilizados para mejorar la fertilidad masculina. Revista Iberoamericana de Fertilidad y Reproducción humana 2014;31(3): 21-25
Boxmeer JC, Smit M, Utomo E, Romijn JC, Eijkemans MJ, Lindemans J et al. Low folate in seminal plasma is associated with increased sperm DNA damage. Fertil Steril. (2009) 92(2):548-556
Cao L, Leers-Sucheta S, Azhar S. Aging alters the functional expression of enzymatic and non-enzymatic anti-oxidant defense systems in testicular rat Leydig cells. J Steroid Biochem Mol Biol. 2004 Jan;88(1):61-7.
Cocuzza M, Sikka SC, Athayde KS, Agarwal A. Clinical relevance of oxidative stress and sperm chromatin damage in male infertility: an evidence based analysis. Int Braz J Urol. 2007 Sep-Oct;33(5):603-21.
Comhaire FH, Christophe A B, Zalata A A, Dhooge W S. Mahmoud A M A, et al. The effects of combined conventional treatment, oral antioxidants and essential fatty acids on sperm biology in subfertile men. Prostaglandins, leukotrienes and essential fatty acids, 2000; 63(3), 159-165.
Cyrus A, Kabir A, Goodarzi D, Moghimi M. The effect of adjuvant vitamin C after varicocele surgery on sperm quality and quantity in infertile men: a double blind placebo controlled clinical trial. Int Braz J Urol (2015) Mar-Apr;41(2):230-8
Dawson EB, Harris WA, Teter MC, Powell LC. Effect of ascorbic acid supplementation on sperm quality of smokers. Fertility and Sterility (1992) 58,1034-1039.
De Lamirande E, Lamothe G. Reactive oxygen-induced reactive oxygen formation during human sperm capacitation. Free Radic Biol Med. 2009 Feb 15;46(4):502-10.
Drevet JR. The antioxidant glutathione peroxidase family and spermatozoa: a complex story. Mol Cell Endocrinol. 2006 May 16;250(1-2):70-9.Ford WC. Regulation of sperm function by reactive oxygen species. Hum Reprod Update. 2004 Sep-Oct;10(5):387-99.
Drummond GR, Cai H, Davis ME, Ramasamy S, Harrison DG. Transcriptional and posttranscriptional regulation of endothelial nitric oxide synthase expression by hydrogen peroxide. Circ Res. 2000 Feb 18;86(3):347-54
Ebisch IM, Pierik FH, De Jong FH, Thomas CM, Steegers-Theunissen RP. Does folic acid and zinc sulphate intervention affect endocrine parameters and sperm. International Journal of Andrology 2006; 29(2):339–45.
Forgiarini A, Masip MA, Gutiérrez SA. ¿Qué es el estrés oxidativo y cómo afecta a la fertilidad masculina? . Reproducción Asistida ORG. 2023.
Fretz PC, Sandlow JI. Varicocele: current concepts in pathophysiology, diagnosis, and treatment. Urol Clin North Am. 2002 Nov;29(4):921-37.
Gandini L, Lombardo F, Paoli D, Caponecchia L, Familiari G, Verlengia C, et al. Study of apoptotic DNA fragmentation in human spermatozoa. Hum Reprod. 2000 Apr;15(4):830-9.
Greco E, Lacobelli M, Rienzi L, Ubaldi F, Susanna F, Tesarik J. Journal of Andrology (2005). Vol 6 3:349-353.
Griveau JF, Dumont E, Renard P, Callegari JP, Le Lannou D. Reactive oxygen species, lipid peroxidation and enzymatic defence systems in human spermatozoa. J Reprod Fertil. 1995 Jan;103(1):17-26.
Griveau JF, Le Lannou D. Reactive oxygen species and human spermatozoa: physiology and pathology. Int J Androl. 1997 Apr;20(2):61-9.
He J, Jin H, Guo J, Li K, Jia L, Li Y, Zhang L. T-2 toxin-induced testicular impairment by triggering oxidative stress and ferroptosis. Ecotoxicol Environ Saf. 2023 Dec 21;270:115844. doi: 10.1016/j.ecoenv.2023.115844. PMID: 38134641.
Homma-Takeda S, Hiraku Y, Ohkuma Y, Oikawa S, Murata M, Ogawa K, et al. 2,4,6-trinitrotoluene-induced reproductive toxicity via oxidative DNA damage by its metabolite. Free Radic Res. 2002 May;36(5):555-66.
Hsu PC., Liu MY, Hsu CC, Chen LY, Guo YL. Lead exposure causes generation of reactive oxygen species and functional impairment in rat sperm. Toxicology. 1997 Sep 26;122(1-2):133-43.
Iwanier K, Zachara BA. Selenium supplementation enhances the element concentration in blood and seminal fluid but does not change the spermatozoa quality characteristics in subfertile men. Journal of Andrology(1995) 16:441-447.
Keske-Ammar L, Feki-Chakroun N, Rebai T, Ghozzi H, Hammami S, Zghal K et al .Sperm oxidative stress and the effect of an oral vitamin e and selenium supplement on semen quality in infertile men. Archives of Andrology (2003) 49:83-94.
Kodama H, Yamaguchi R, Fukuda J, Kasai H, Tanaka T. Increased oxidative deoxyribonucleic acid damage in the spermatozoa of infertile male patients. Fertility and Sterility (1997) 68,519-524.
Ko EY, Sabanegh ES Jr, Agarwal A3. Male infertility testing: reactive oxygen species and antioxidant capacity. Fertil Steril. 2014 Dec;102(6):1518-27.
Koizumi T, Li ZG. Role of oxidative stress in single-dose, cadmium-induced testicular cancer. J Toxicol Environ Health. 1992 Sep;37(1):25-36.
Kothari S, Thompson A, Agarwal A, du Plessis SS. Free radicals: their beneficial and detrimental effects on sperm function. Indian J Exp Biol. 2010 May;48(5):425-35.
La OMS alerta de que una de cada seis personas padece esterilidad. Who.int.
Lenzi A, Lombardo F, Sgró P, Salacone P, Caponecchia L, Dondero F et al . Use of carnitine therapy in selected cases of male factor infertility: a double-blind crossover trial. Fertility and Sterility (2003), 79(2): 292–300
Lewis SE, Sterling ES, Young IS, Thompson. Comparison of individual antioxidants of sperm and seminal plasma in fertile and infertile men. Fertility and Sterility (1997) 67,142-147.
Lifeng Yan, Jining Liu, Shengmin Wu, Shenghu Zhang, Guixiang Ji, Aihua Gu. Seminal superoxide dismutase activity and its relationship with semen quality and SOD gene polymorphism. J Assist Reprod Genet. 2014 May; 31(5): 549–554.
Li YC, Hu XQ, Xiao LJ, Hu ZY, Guo J, Zhang KY, et al. An oligonucleotide microarray study on gene expression profile in mouse testis of experimental cryptorchidism. Front Biosci. 2006 Sep 1;11:2465-82.
López G, Martínez Soto JC, Amorós D. Factor masculino y estrés oxidativo. Cuadernos de Andrología Clínica: Contaminación ambiental y manejo del estrés oxidativo en el factor masculino. 2017;(Asebir):47–53.
Makker K, Agarwal A, Sharma R. Oxidative stress & male infertility. Indian J Med Res. 2009 Apr;129(4):357-67.
Marchlewicz M, Wiszniewska B, Gonet B, Baranowska-Bosiacka I, Safranow K, Kolasa A, et al. Increased lipid peroxidation and ascorbic Acid utilization in testis and epididymis of rats chronically exposed to lead. Biometals. 2007 Feb;20(1):13-9.
Martínez Soto JC, Amorós D, López G. Sistema antioxidante en semen y suplementación exógena. Cuadernos de Andrología Clínica: Contaminación ambiental y manejo del estrés oxidativo en el factor masculino. 2017;(Asebir):54–64.
Mattison DR The effects of smoking on fertility from gametogenesis to implantation. Environ Res. 1982 Aug;28(2):410-33.
Morgante G, Scolaro V, Tosto C, Di Sabatino A, Piomboni P, De Leo V. Treatment with carnitine, acetyl carnitine, Larginine and ginseng improves sperm motility and sexual health in men with asthenopermia. Minerva Urologica e Nefrologica 2010;62(3):213–218
Moustafa MH, Sharma RK, Thornton J, Mascha E, Abdel-Hafez MA, Thomas AJ Jr, et al. Relationship between ROS production, apoptosis and DNA denaturation in spermatozoa from patients examined for infertility. Hum Reprod. 2004 Jan;19(1):129-38.
Nicolich A, Lafuente R, López G, García-Peiró A, Brassesco M. Análisis del estrés oxidativo en el eyaculado mediante la determinación del anión superóxido. Revista Asebir, el 1 de junio de 2013;18 (1)(Junio 2013):15–19.
Omu AE, Al Azemi MK, Kehinde EO, Anim JT, Oriowi MA. Mathew TC. Indications of the mechanisms involved in improved sperm parameters by zinc therapy. Medical principes and practice (2008) 17(2):108-116
Omu AE, Dashtib H,Othmanc SA. Treatment of asthenozoospermia with zinc sulphate: andrological, immunological and obstetric outcome. European Journal of Obstetrics & Gynecology and Reproductive Biology (1998) ,79(2):179-184.
O›Flaherty C1, de Souza AR. Hydrogen peroxide modifies human sperm peroxiredoxins in a dose-dependent manner. Biol Reprod. 2011 Feb;84(2):238-47.
Rayman, Margaret P. The importance of selenium to human health. The Lancet (2000) , Vol 356 , 9225: 233 - 241
Reddy MM, Mahipal SV, Subhashini J, Reddy MC, Roy KR, Reddy GV, et al. Bacterial lipopolysaccharide-induced oxidative stress in the impairment of steroidogenesis and spermatogenesis in rats. Reprod Toxicol. 2006 Oct;22(3):493-500.
Safarinejad M R, Hosseini S Y, Dadkhah F, Asgari M A. Relationship of omega-3 and omega-6 fatty acids with semen characteristics, and anti-oxidant status of seminal plasma: A comparison between fertile and infertile men. Clinical Nutrition 2010 29, 100-105.
Safarinejad M R. Effect of omega-3 polyunsaturated fatty acid supplementation on semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia 2011; 43, 38-47.
Safarinejad MR. Efficacy of coenzyme Q10 on semen parameters, sperm function and reproductive hormones in infertile men. J Urol (2009);182:237-248
Scott R, Macpherson A, Yates RW, Hussain, Dixon. The effect of selenium supplementation on human sperm motility. British journal of Urology (1998) 82:76-80.
Sharma RK, Agarwal A. Role of reactive oxygen species in male infertility. Urology. 1996 Dec;48(6):835-50.
Sikka SC. Role of oxidative stress and antioxidants in andrology and assisted reproductive technology. J Androl. 2004 Jan-Feb;25(1):5-18.
Suleiman SA, Ali ME, Zaki ZM, El-Malik EMA, Nasr MA . Lipid peroxidation and human sperm motility: protective role of vitamin E. Journal of Andrology (1996)17:530-537.
Syed V, Hecht NB. Rat pachytene spermatocytes down-regulate a polo-like kinase and up-regulate a thiol-specific antioxidant protein, whereas sertoli cells down-regulate a phosphodiesterase and up-regulate an oxidative stress protein after exposure to methoxyethanol and methoxyacetic acid. Endocrinology. 1998 Aug;139(8):3503-11.
Tremellen K. Oxidative stress and male infertility--a clinical perspective. Hum Reprod Update. 2008 May-Jun;14(3):243-58.
Vernet P1, Aitken RJ, Drevet JR. Antioxidant strategies in the epididymis. Mol Cell Endocrinol. 2004 Mar 15;216(1-2):31-9.
Vicari E and Calogero AE. Effects of treatment with carnitines in infertile patients with prostato-vesiculo-epididymitis. Human Reproduction (2001) 16,2338-2342.
Yu B, Huang Z. Variations in Antioxidant Genes and Male Infertility. Biomed Res Int. 2015;2015:513196
Está muy bien, con una gran cantidad de información y referencias. Dadle un repaso, ya que la expresión se puede mejorar. La estructura hace la redacción algo repetitiva; sobre todo, evitad párrafos demasiado cortos o de una sola frase.
ResponderEliminarRevisad los pequeños errores de expresión o formato (espaciado, algún paréntesis); revisad súper/subíndices, es importante escribir las fórmulas y términos correctamente. Los nombres de enzimas y moléculas no deberían ir capitalizados, son nombres comunes (a menos que lleven un nombre propio).
La tabla final debería estar citada en el texto.
EliminarTened cuidado con declaraciones como la frase final "La L-arginina es un aminoácido cuya administración combinada con carnitina y ginseng durante tres meses mejora la movilidad espermática". En primer lugar, es un trabajo aislado, considerad estos resultados críticamente. De hecho, creo que queríais poner algo sobre cada antioxidante, pero tal vez no sea lo más apropiado. Deberíais escribir en general para cada punto (mecanismo, qué beneficio celular resulta), y, si procede, un ejemplo (La L-arginina tiene actividad antioxidante... Por ejemplo, Morgante et al. (2010) indicaron que podría beneficiar... en un tratamiento combinado con...).
¿El resumen gráfico es de producción propia?
El gráfico si es de producción propia y tiene razón en cuanto a la citación de la tabla, deberíamos haberla citado en el texto y enlazarla con el argumento. Por otra parte, tendremos en cuenta su crítica para futuras redacciones en cuanto a la expresión y estructura. Ajeno a ello, esperamos que le haya gustado y le resulte interesante el tema elegido.
ResponderEliminarRecordad que esta fase es para revisar los dos trabajos. He realizado una primera evaluación, pero puede mejorar.
ResponderEliminarYa hemos realizado los cambios indicados. Como ha indicado Estrella, tanto el resumen gráfico como las figuras del otro artículo son de elaboración propia.
ResponderEliminarMuy bien. Sobre el resumen gráfico, enhorabuena, es una buena idea (por eso os preguntaba).
ResponderEliminar